

What Is The Chemical Makeup For Rna

A hairpin loop from a pre-mRNA. Notice the single strand with its nitrogen-rich (bluish) bases extending from its oxygen-rich (red) backbone.
Ribonucleic acrid or RNA is a polymer or chain of nucleotide units, each comprising a nitrogenous base (adenine, cytosine, guanine, or uracil), a five-carbon sugar (ribose), and a phosphate group. The sugar and phosphate groups class the polymer's backbone, while the nitrogenous bases extending from the backbone provide RNA'due south distinctive properties.
In living cells, RNA in different configurations fulfills several of import roles in the process of translating genetic information from dna (Dna) into proteins. One type of RNA (messenger(m) RNA) acts as a messenger between Deoxyribonucleic acid and the protein synthesis complexes known equally ribosomes; a 2d blazon (ribosomal(r) RNA) forms vital portions of the structure of ribosomes; a third blazon (transfer(t) RNA) is an essential guide to deliver the appropriate poly peptide building blocks, amino acids, to the ribosome; and other types of RNA, microRNAs (miRNAs) play a part in regulating gene expression, while minor nuclear(sn) RNA helps with assuring that mRNA contains no nucleotide units that would atomic number 82 to formation of a faulty protein. RNA too serves equally a genetic blueprint for certain viruses, and some RNA molecules (chosen ribozymes) are likewise involved in the catalysis of biochemical reactions.
RNA is very like to DNA, just differs in a few important structural details. RNA is unremarkably single stranded, while DNA naturally seeks its stable form as a double stranded molecule. RNA nucleotides contain ribose while Deoxyribonucleic acid nucleotides contain the closely related sugar deoxyribose. Furthermore, RNA uses the nucleotide uracil in its limerick, instead of the thymine that is present in DNA. RNA is transcribed from Dna by enzymes called RNA polymerases and is by and large farther candy by other enzymes, some of them guided past non-coding RNAs.
Single-stranded RNA is like to the protein polymer in its natural propensity to fold back and double up with itself in complex means assuming a multifariousness of biologically useful configurations.
Contents
- ane Chemical and stereochemical construction
- 2 Comparison with DNA
- 3 Synthesis
- 4 Biological roles
- four.i Messenger RNA (mRNA)
- 4.ii Non-coding RNA
- 4.2.1 Transfer RNA (tRNA)
- 4.2.2 Ribosomal RNA (rRNA)
- four.2.3 Catalytic RNA
- four.three Genetic blueprint in some viruses
- 5 RNA world hypothesis
- 6 RNA secondary structures
- 7 History
- 8 List of RNA types
- 9 References
- x External links
- 11 Credits
The connectedness of living organisms can be seen in the ubiquitousness of RNA in living cells and in viruses throughout nature, and in the universal role of RNA in protein synthesis.
Chemical and stereochemical structure
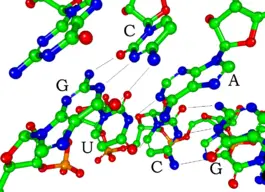
Base of operations-pairing in a siRNA (small interfering RNA) segment, a double-stranded type of RNA. Hydrogen atoms are not shown.
RNA is a nucleic acrid, a complex, loftier-molecular-weight macromolecule composed of nucleotide chains whose sequence of bases conveys genetic data.
A nucleotide is a chemic compound comprising three components: a nitrogen-containing base, a pentose (five-carbon) sugar, and ane or more than phosphate groups. The nitrogen-containing base of a nucleotide (too chosen the nucleobase) is typically a derivative of either purine or pyrimidine. The most common nucleotide bases are the purines adenine and guanine and the pyrimidines cytosine and thymine (or uracil in RNA).
Nucleic acids are polymers of repeating units (chosen monomers). Specifically, they oft incorporate long chains of nucleotide monomers connected by covalent chemical bonds. RNA molecules may comprise as few as 75 nucleotides or more than v,000 nucleotides, while a Deoxyribonucleic acid molecule may comprise more than 1,000,000 nucleotide units.

A conventional skeletal formula
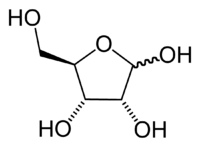
In RNA, the sugar component, ribose is a h2o-soluable, pentose sugar (monosaccharide with five carbon atoms). Ribose has the chemical formula CfiveHtenO5.
Ribose is an aldopentose, which means a pentose sugar with an aldehyde functional group in position one. An aldehyde group comprises a carbon atom bonded to a hydrogen atom and double-bonded to an oxygen atom (chemical formula O=CH-). Ribose forms a 5-member ring with four carbon atoms and one oxygen. Hydroxyl (-OH) groups are fastened to iii of the carbons. The fourth carbon in the ring (ane of the carbon atoms adjacent to the oxygen) has attached to it the fifth carbon atom and a hydroxyl group.
The RNA polymer features a ribose and phosphate courage with one of 4 different nucleotide bases—adenine, guanine, cytosine, and uracil—fastened to each ribose-phosphate unit.
At that place are also numerous modified bases and sugars establish in RNA that serve many different roles. Pseudouridine (Ψ), in which the linkage between uracil and ribose is changed from a C–N bond to a C–C bail, and ribothymidine (T), are institute in various places (nearly notably in the TΨC loop of tRNA). Another notable modified base is hypoxanthine (a deaminated guanine base of operations whose nucleoside is called inosine). Inosine plays a cardinal function in the Wobble Hypothesis of the genetic code. There are nearly 100 other naturally occurring modified nucleosides, of which pseudouridine and nucleosides with two'-O-methylribose are by far the about common. The specific roles of many of these modifications in RNA are not fully understood. Even so, information technology is notable that in ribosomal RNA, many of the post-translational modifications occur in highly functional regions, such as the peptidyl transferase eye and the subunit interface, implying that they are important for normal function.
The most important structural feature of RNA that distinguishes it from DNA is the presence of a hydroxyl group at the ii'-position of the ribose sugar. The presence of this functional group enforces the C3'-endo saccharide conformation (as opposed to the C2'-endo conformation of the deoxyribose sugar in Dna) that causes the helix to prefer the A-class geometry rather than the B-grade most normally observed in Dna. This results in a very deep and narrow major groove and a shallow and wide minor groove. A second consequence of the presence of the two'-hydroxyl group is that in conformationally flexible regions of an RNA molecule (that is, not involved in germination of a double helix), it tin chemically attack the adjacent phosphodiester bail to cleave the courage.
Comparison with Dna

Left: An RNA strand, with its nitrogenous bases. Right: Double-stranded Deoxyribonucleic acid.
The well-nigh common nucleic acids are deoxyribonucleic acrid (Deoxyribonucleic acid) and ribonucleic acid (RNA). The primary role of DNA is the long-term storage of genetic data. Dna is oftentimes compared to a pattern, since it contains instructions for constructing other components of the cell, such as proteins and RNA molecules. The DNA segments that acquit genetic information are called genes, but other Dna sequences have structural purposes or are involved in regulating the expression of genetic information. RNA, also, may serve more than than one purpose, just it is most ordinarily identified as the intermediate betwixt the DNA blueprint and the actual workings of the cell, serving every bit the template for the synthesis of proteins from the genetic information stored in Deoxyribonucleic acid.
RNA and Deoxyribonucleic acid differ in three main ways.
First, unlike Deoxyribonucleic acid which is double-stranded, RNA is intrinsically a single-stranded molecule in most of its biological roles and has a much shorter chain of nucleotides. (While RNA is usually unmarried-stranded, the RNA molecule also quite commonly forms double-helical regions where a given strand has folded dorsum on itself. Double-stranded RNA is constitute likewise in certain viruses.)
Secondly, while DNA contains deoxyribose, RNA contains ribose. In that location is no hydroxyl group attached to the pentose ring in the 2' position in DNA, whereas RNA has ii hydroxyl groups. These hydroxyl groups make RNA less stable than Dna considering it is more prone to hydrolysis. ("Deoxy" simply indicates that the sugar lacks an oxygen atom present in ribose, the parent compound.)
Thirdly, the complementary nucleotide to adenine is not thymine, as it is in DNA, merely rather uracil, which is an unmethylated form of thymine.
Most biologically active RNAs, including tRNA, rRNA, snRNAs, and other non-coding RNAs (such as the point recognition particle(SRP) RNAs), comprise extensively base of operations paired regions that have folded together to form double stranded helices. Structural analysis of these RNAs reveals that they are highly structured with tremendous variety with collections of short helices packed together into structures much more than akin to proteins than to DNA, which is unremarkably limited to long double-stranded helices. Through such a variety of structures, RNAs can achieve chemical catalysis, like enzymes. For instance, determination of the structure of the ribosome—an enzyme that catalyzes peptide bail formation—revealed that its active site is composed entirely of RNA.
Synthesis
Synthesis of RNA is usually catalyzed by an enzyme, RNA polymerase, using Deoxyribonucleic acid as a template. Initiation of synthesis begins with the binding of the enzyme to a promoter sequence in the Dna (usually found "upstream" of a cistron). The Dna double helix is unwound by the helicase activity of the enzyme. The enzyme then progresses along the template strand in the iii' -> 5' management, synthesizing a complementary RNA molecule with elongation occurring in the 5' -> 3' direction. The DNA sequence also dictates where termination of RNA synthesis will occur (Nudler and Gottesman 2002).
There are also a number of RNA-dependent RNA polymerases as well that utilize RNA every bit their template for synthesis of a new strand of RNA. For example, a number of RNA viruses (such as poliovirus) use this type of enzyme to replicate their genetic material (Hansen et al. 1997). Also, it is known that RNA-dependent RNA polymerases are required for the RNA interference pathway in many organisms (Ahlquist 2002).
Biological roles
RNA's slap-up variety of possible structures and chemical backdrop permits it to perform a much greater diverseness of roles than in the cell than Deoxyribonucleic acid. Three main types of RNA are involved in protein synthesis:
- Messenger RNA (mRNA) serves equally the template for the synthesis of a protein. It carries data from Deoxyribonucleic acid to the ribosome.
- Transfer RNA (tRNA) is a small chain of nucleotides that transfers a specific amino acrid to a growing polypeptide chain at the ribosomal site of synthesis. Information technology pairs the amino acid to the appropriate three-nucleotide codon on the mRNA molecule.
- Ribosomal RNA (rRNA) molecules are extremely abundant and make upwardly at to the lowest degree 80 percentage of the RNA molecules found in a typical eukaryotic jail cell. In the cytoplasm, usually three or 4 rRNA molecules combine with many proteins to perform a structural and essential catalytic role, as components of the ribosome.
RNA also may serve every bit a catalyst for reactions and as a genetic blueprint, rather than DNA, in various viruses. Some RNA, including tRNA and rRNA, is non-coding in that information technology is not translated into proteins.
Messenger RNA (mRNA)
Messenger RNA is RNA that carries data from DNA to the ribosome sites of poly peptide synthesis in the cell. In eukaryotic cells, once mRNA has been transcribed from DNA, it is "processed" earlier being exported from the nucleus into the cytoplasm, where information technology is spring to ribosomes and translated into its corresponding protein form with the help of tRNA. In prokaryotic cells, which practice non take nucleus and cytoplasm compartments, mRNA tin can bind to ribosomes while it is being transcribed from Deoxyribonucleic acid. Later on a certain amount of time the message degrades into its component nucleotides, usually with the assistance of ribonucleases.
Non-coding RNA
RNA genes (also known as non-coding RNA or small RNA) are genes that encode RNA that is not translated into a poly peptide. The most prominent examples of RNA genes are those coding for transfer RNA (tRNA) and ribosomal RNA (rRNA), both of which are involved in the process of translation. Two other groups of non-coding RNA are microRNAs (miRNA) which regulate the expression of genes through a procedure called RNA interference (RNAi), and minor nuclear RNAs (snRNA), a diverse class that includes for example the RNAs that class spliceosomes that excise introns from pre-mRNA (Berg et al. 2002).
Transfer RNA (tRNA)
Transfer RNA is a pocket-size RNA chain of about 74-95 nucleotides that transfers a specific amino acid to a growing polypeptide concatenation at the ribosomal site of protein synthesis, during translation. Information technology has sites for amino-acrid attachment and an anticodon region for codon recognition that binds to a specific sequence on the messenger RNA chain through hydrogen bonding. It is a type of non-coding RNA.
Ribosomal RNA (rRNA)
Ribosomal RNA is the catalytic component of the ribosomes, the poly peptide synthesis factories in the cell. Eukaryotic ribosomes comprise 4 different rRNA molecules: 18S, 5.8S, 28S, and 5S rRNA. Three of the rRNA molecules are synthesized in the nucleolus, and one is synthesized elsewhere. rRNA molecules are extremely abundant and make up at least 80 percent of the RNA molecules found in a typical eukaryotic cell.
Catalytic RNA
Although RNA contains only iv bases, in comparison to the twenty-odd amino acids commonly found in proteins, certain RNAs (called ribozymes) are nevertheless able to catalyze chemical reactions. These include cutting and ligating other RNA molecules, and also the catalysis of peptide bond germination in the ribosome.
Genetic blueprint in some viruses
Some viruses contain either single-stranded or double-stranded RNA equally their source of genetic data. Retroviruses, for example, store their genetic data every bit RNA, though they replicate in their hosts via a DNA intermediate. Once in the host's cell, the RNA strands undergo contrary transcription to DNA in the cytosol and are integrated into the host'due south genome. Man immunodeficiency virus (or HIV) is a retrovirus thought to cause acquired immune deficiency syndrome (AIDS), a condition in which the human immune system begins to neglect, leading to life-threatening opportunistic infections.
Double-stranded RNA (dsRNA) is RNA with two complementary strands, similar to the Dna found in all cells. dsRNA forms the genetic material of some viruses chosen dsRNA viruses. In eukaryotes, long RNA such every bit viral RNA can trigger RNA interference, where short dsRNA molecules called siRNAs (small interfering RNAs) can cause enzymes to break downwardly specific mRNAs or silence the expression of genes. siRNA can also increase the transcription of a gene, a process chosen RNA activation (Doran 2007). siRNA is often confused with miRNA; siRNAs are double-stranded, whereas miRNAs are unmarried-stranded.
RNA world hypothesis
The RNA world hypothesis proposes that the primeval forms of life relied on RNA both to carry genetic information (like Dna does now) and to catalyze biochemical reactions similar an enzyme. According to this hypothesis, descendants of these early lifeforms gradually integrated DNA and proteins into their metabolism.
In the 1980s, scientists discovered that certain RNA molecules (called ribozymes) may office as enzymes, whereas previously merely proteins were believed to have catalytic power. Many natural ribozymes catalyze either their ain cleavage or the cleavage of other RNAs, but they take also been found to catalyze the aminotransferase action of the ribosome.
The discovery of ribozymes provides a possible explanation for how early RNA molecules might have get-go catalyzed their own replication and developed a range of enzymatic activities. Known as the RNA world hypothesis, this explanation posits that RNA evolved before either DNA or proteins from free-floating nucleotides in the early "primordial soup." In their part as enzymes, RNA molecules might have begun to catalyze the synthesis of proteins, which are more than versatile than RNA, from amino acrid molecules. Next, DNA might have been formed by reverse transcription of RNA, with DNA eventually replacing RNA as the storage form of genetic textile. Although at that place are remaining difficulties with the RNA world hypothesis, it remains as a possible key to agreement the origin and development of the multi-functional nature of nucleic acids, the interconnectedness of life, and its common origins.
RNA secondary structures
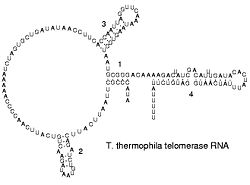
Secondary structure of an RNA from a telomerase.
The functional grade of single stranded RNA molecules, just like proteins, frequently requires a specific tertiary structure. The scaffold for this structure is provided by secondary structural elements, which arise through the formation of hydrogen bonds within the interfolded molecule. This leads to several recognizable "domains" of secondary construction like hairpin loops, bulges, and internal loops. The secondary structure of RNA molecules tin can be predicted computationally by calculating the minimum free energies (MFE) structure for all different combinations of hydrogen bondings and domains (Mathews et al. 2004). There has been a significant amount of research directed at the RNA structure prediction problem.
History
Nucleic acids were discovered in 1868 past Johann Friedrich Miescher (1844-1895), who called the material 'nuclein' since information technology was plant in the nucleus. Information technology was afterwards discovered that prokaryotic cells, which exercise not have a nucleus, too contain nucleic acids.
The role of RNA in poly peptide synthesis had been suspected since 1939, based on experiments carried out by Torbjörn Caspersson, Jean Brachet, and Jack Schultz. Hubert Chantrenne elucidated the messenger function played by RNA in the synthesis of proteins in ribosomes. Finally, Severo Ochoa discovered RNA, winning Ochoa the 1959 Nobel Prize for Medicine. The sequence of the 77 nucleotides of a yeast RNA was found by Robert W. Holley in 1964, winning Holley the 1968 Nobel Prize for Medicine. In 1976, Walter Fiers and his team at the University of Ghent determined the complete nucleotide sequence of bacteriophage MS2-RNA (Fiers et al. 1976).
List of RNA types
| Type | Function | Distribution |
|---|---|---|
| mRNA | Codes for protein | All cells |
| rRNA | Translation | All cells |
| tRNA | Translation | All cells |
| snRNA | RNA modification | All cells |
| snoRNA | RNA modification | All cells |
| miRNA | Gene regulation | Eukaryotes |
| piRNA | Gene regulation | Brute germline cells |
| siRNA | Gene regulation | Eukaryotes |
| Antisense mRNA | Preventing translation | Bacteria |
| tmRNA | Terminating translation | Bacteria |
| SRP RNA | mRNA tagging for export | All cells |
| Ribozyme | Catalysis | All cells |
| Transposon | Cocky-propagating | All cells |
| Viroid | Cocky-propagating | Infected plants |
In addition, the genome of many types of viruses consists of RNA, namely:
- Double-stranded RNA viruses
- Positive-sense RNA viruses
- Negative-sense RNA viruses
- Retroviruses
- Satellite viruses
References
ISBN links support NWE through referral fees
- Ahlquist, P. 2002. RNA-dependent RNA polymerases, viruses, and RNA silencing. Science 296(5571): 1270-1273.
- Berg, J. M., J. L. Tymoczko, and Fifty. Stryer. 2002. Biochemistry, 5th Edition. WH Freeman and Company. ISBN 0716746840.
- Doran, Grand. 2007. RNAi – Is one suffix sufficient? Periodical of RNAi and Gene Silencing 3(i): 217-219. Retrieved December 7, 2007.
- Fiers W et al. 1976. Complete nucleotide-sequence of bacteriophage MS2-RNA: Master and secondary structure of replicase gene. Nature 260: 500-507.
- Hansen, J. L., A. K. Long, and Due south. C. Schultz. 1997. Structure of the RNA-dependent RNA polymerase of poliovirus. Structure five(8): 1109-1122. Retrieved December 7, 2007.
- Mathews, D. H., Chiliad. D. Disney, J. L. Childs, S. J. Schroeder, G. Zuker, and D. H. Turner. 2004. Incorporating chemic modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proc. Natl. Acad. Sci. U. Southward. A. 101(19): 7287-7292. Retrieved December half dozen, 2007.
- Nudler, E., and Thou. E. Gottesman. 2002. Transcription termination and anti-termination in E. coli. Genes to Cells 7: 755-768. Retrieved December 7, 2007.
External links
All links retrieved June 17, 2019.
- RNAJunction Database: Extracted atomic models of RNA junction and kissing loop structures.
Credits
New World Encyclopedia writers and editors rewrote and completed the Wikipedia article in accordance with New World Encyclopedia standards. This commodity abides by terms of the Creative Commons CC-by-sa iii.0 License (CC-by-sa), which may exist used and disseminated with proper attribution. Credit is due under the terms of this license that can reference both the New Earth Encyclopedia contributors and the selfless volunteer contributors of the Wikimedia Foundation. To cite this article click here for a list of acceptable citing formats.The history of before contributions by wikipedians is accessible to researchers here:
- RNA history
The history of this article since it was imported to New Earth Encyclopedia:
- History of "RNA"
Note: Some restrictions may apply to apply of individual images which are separately licensed.
Source: https://www.newworldencyclopedia.org/entry/RNA
Posted by: stewartfortalwyneho.blogspot.com
0 Response to "What Is The Chemical Makeup For Rna"
Post a Comment